咖啡叶锈病:过去,现在和未来(二)
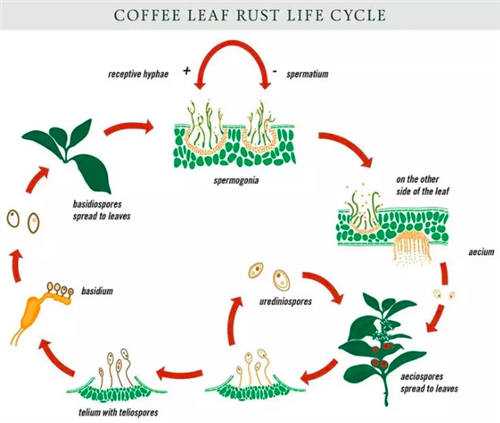
两种不同交配型担孢子(basidospores)单倍体菌丝体配合(spermogonia)----在叶片背面发育成锈孢子器和(aecium )----锈子器成熟后,锈孢子(aeciospores)随风传播到咖啡树由叶子裂缝侵入萌发产生发芽管(咖啡叶上的病斑)----在病斑处形成夏孢子/夏孢子堆(urediniospores)----夏孢子循环侵入咖啡叶----夏孢子转变为冬孢子(telium)----冬孢子萌发担子(basidium)--担孢子(basidospores)---担孢子(basidospores)随风散到转主寄主上侵染.
症状
啡锈病菌按系统分类属于担子菌亚 冬泡菌纲 , 锈菌 目, 柄锈菌科.
最初症状是在咖啡叶背出现一些最大直径为1~ 1.5毫米的淡黄色小圆斑点 ,病斑数取决于侵染程度. 在咖啡叶的叶面相应出现一些透明的侵染斑点,叶片褪色是因海绵薄壁组织细胞内的叶绿体的变化而引起的.
在叶片背面 , 这些斑点的直径在短时间内扩大到3毫米. 在此阶段, 有淡黄色粉状物析出, 随着病程的进展, 在咖啡叶片上出现橙黄锈色. 这就是病菌繁殖器官, 即病原菌匀夏孢子.
叶面象叶背一样, 在斑点周围可见到黄绿色的模糊晕轮. 这些斑点的直径可以扩展到2~ 3厘米. 在一个叶片上能产生 70 多个病斑或疤状突起 , 许多斑点交错排列并完全覆盖住叶片 .
在严重感病的情况下每片叶有2~10个疤状突起 ,严重发病会使咖啡叶片干枯 , 过早落叶直至整个分枝死亡,果实变黑并脱落.
咖啡锈病菌能产生典型肾型多瘤夏孢子(urediniospores) , 夏孢子的大小因种,培养的生理型以及宿主植物的种类而显著不同 . 平均为30~36 x 21~23微米.
如果落在合适的咖啡植株上, 并处于3一24°C的最佳温度且有水滴时 , 夏孢子可在 6小时内萌发,在水蒸汽饱和的空气中萌发迟 , 相对空气湿度为95%时 , 夏孢子不再萌发. 由于这种原因,在海拔较高早地区的咖啡园较少遭受锈病的危害.
咖啡锈病菌能产生一个或多个发芽管,它们不能直接侵入叶片表面 , 而是在叶片表面分枝, 直到遇列裂口时才侵入叶片 .此时 , 病原茵就能形成一令附着孢 , 刺入菌丝由此处侵入到宿主植株的气下室 , 形成一个气囊并以吸器从宿主细胞内摄取营养 .
咖啡锈病菌在海绵组织中的细胞间蔓延并发育成一个具有大量吸器的多分枝菌丝体. 从个别咖啡品种感病性看, 相当宽的菌丝( 6微米)能深浅不同地侵入到栅栏薄壁组织内.
咖啡锈病菌从开怡侵染到生第一个孢子之间的平均时间为4-7周,取于气候条件 , 叶龄 , 感病部位 , 栽培品种或无性系的敏感性.夏孢子能重新侵染新叶 , 这个过 程在生长周期中常常频繁地重复.
关于风和雨在夏孢子传播中的作用有不同的观点 .病害在亚洲南部和东部国家里迅速传播似乎证明风对夏孢子的传播有着重要意义.肯尼亚的比较试验表明, 风是夏孢子的主要传播媒介.夏孢子的弱点是其遗传的单一性和对低温和干燥天气的敏感性.
锈菌具有内置的生物保护机制,可以抵御寒冷、干旱气候.在这些条件下相同的夏孢子转变为陀螺形的子孢子, 也就是冬孢子(teliospore).冬孢子是锈菌的休眠和越冬形态.
冬孢子萌发出一个较长菌丝 , 菌丝横断并形成球形担孢子(basidospores).然而 , 这些担抱子不能侵入咖啡叶组织. 由于这一原因; 有人推测咖啡锈病必须经风媒寻找并通过其他宿主植物来完成锈病的发育过程 .
锈菌这种双寄主关系在小麦和浆果上得到了很好的证明.然而,咖啡生长在炎热潮湿的热带地区,阿拉比卡几乎不能忍受霜冻,更不用说冰冻的低温度.因此,咖啡锈菌(Hemileia vastatrix)甚至不需要第二个宿主.事实上,咖啡叶锈病很容易以夏孢子传播开,科学目前尚未发现确定中间宿主. 有一点显然可以肯定 , 即锈病菌不能在咖啡树群中完成其完整的生活周期.
想一下.咖啡叶锈病是如此的复杂,它甚至不需要完成它的生命周期就可以毁灭一个咖啡农场.这暗示了一个巨大问题:锈菌在"学习"如何适应环境,在未来很长一段时间内会继续以咖啡为食.
标签: 咖啡叶锈病
上一篇:咖啡叶锈病:过去,现在和未来
下一篇:咖啡叶锈病:过去,现在和未来(三)